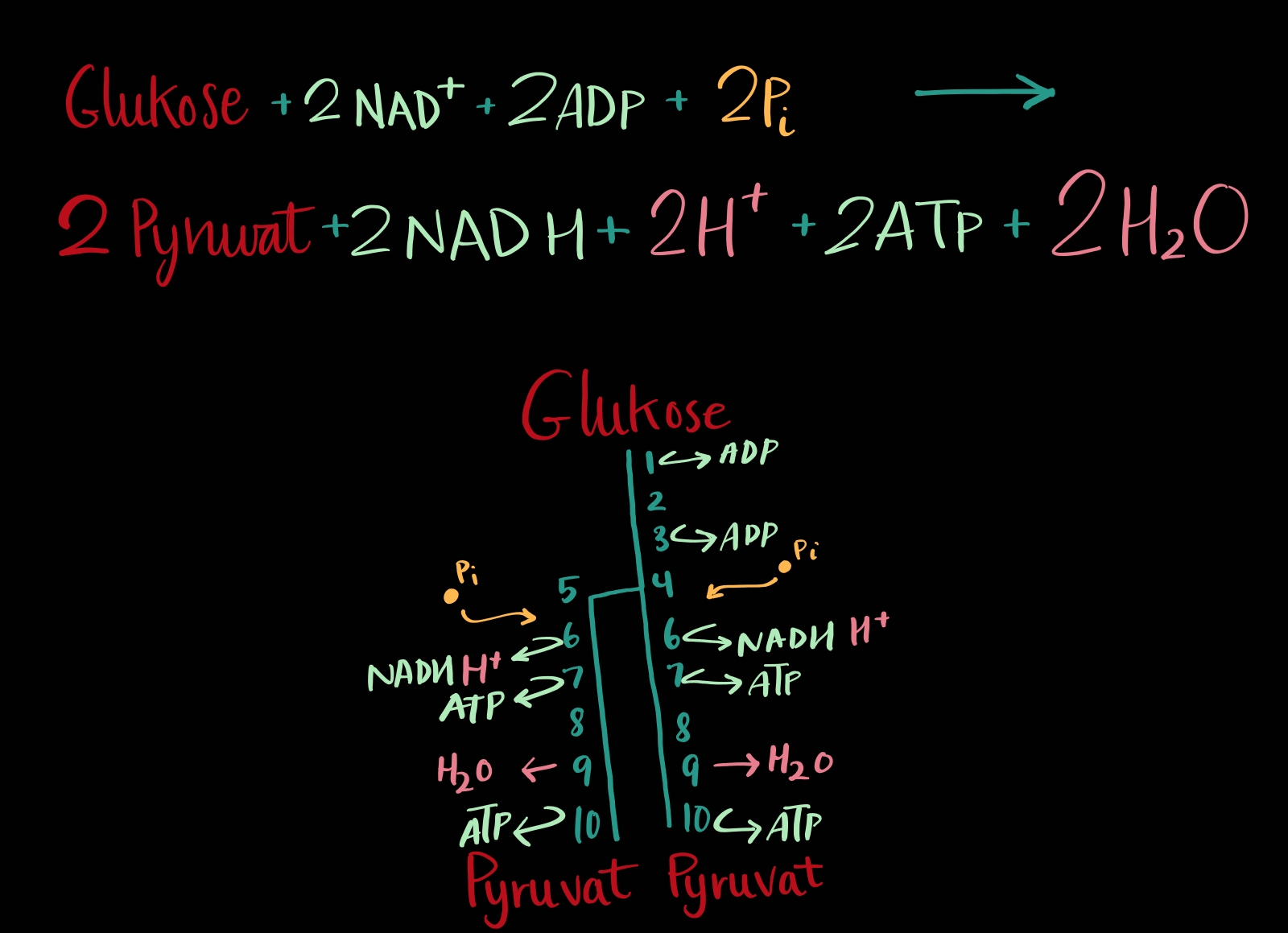
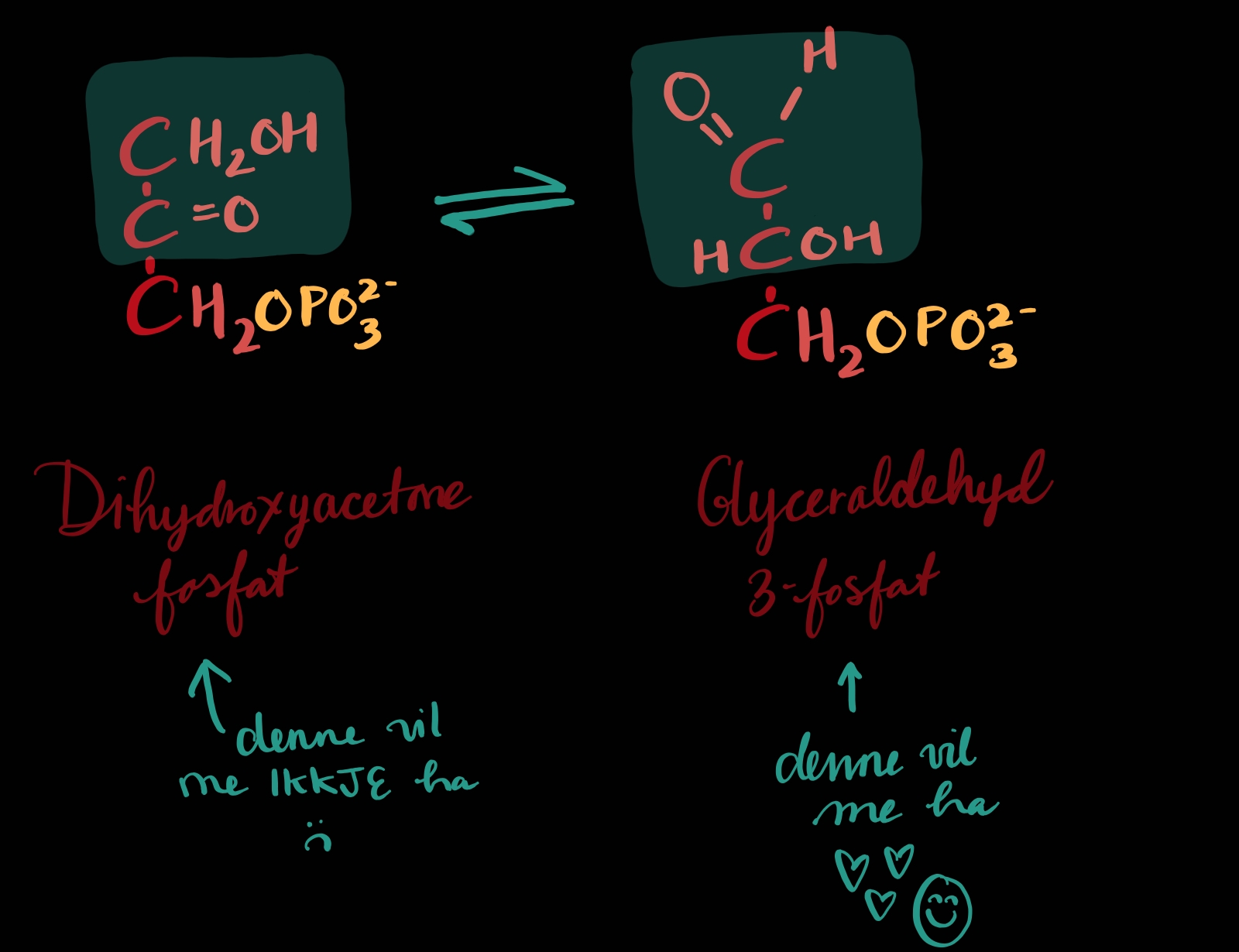Eller sukkersplitting/søtsplitting, om du vil, er når eit sukkermolekyl, glukose, blir delt i to mindre molekyl, pyruvat. Glukosen har seks karbon og dei to pyruvata har tre karbon kvar. Dette er ein prosess som produserer bittelitt 🤏 energi, men for lite for dei fleste celler å nøye seg med. Glykolysen er fyrste del av celleånding, og i løpet av heile celleåndinga så blir det produsert nok energi for cellene å leve på (men ein kan ikkje hoppe over noko steg, og glykosen er kjempeviktig sjølv om energiproduksjonen er liten, kvifor? Fordi det er steg1).
Kjært barn har mange namn og eg blir lett forvirra. Derfor er eg IKKJE konsekvent når det gjeld alle benevningar særleg fosfat og fosforyl. Forskjellen er berre eit oksygen, og det er ikkje så viktig i denne samanhengen her spør du meg. Andre kjære barn er sjølve glykoslysen så her er ulike måtar å skildra han på: Glykolyse er nedbryting[1], glykolyse er oksidasjon[2] glykolyse er katabolisme[3].
Glykolysen er ein ti-stegs prosess som i mine bøker blir skildra som ein vertikal strek, men eg synst han er betra teikna som ein stemmegaffel slik eg har gjort på min teikning. Molekylet blir nemleg delt i to på trinn4, så trinn6 til 10 blir gjort to gonger for kvart glukosemolekyl. Trinn5 blir berre gjort ein gong, for dei to halvdelane frå trinn4 er ikkje like så då blir den eine endra på i trinn5. Alle dei ti stega bruker kvart sitt unike enzym, altså er det ingen trinn som må slost om enzyma – alle har sitt eige.
PS: Enn så lenge kan alle bildene høgreklikkast og opnast i ny fane og blir kjempestore, fordi eg hadde ikkje gidda til å gjere dei til passeleg storleik. Dette kan endre seg.
2: Glukose blir oksidert og me får NADH og ATP. Oksidasjon er tap av elektron.
3: Katabolisme er eit fint ord for å bryte ned (ned+kaste) og glukose blir altså katabolisert/nedbrutt til pyruvat
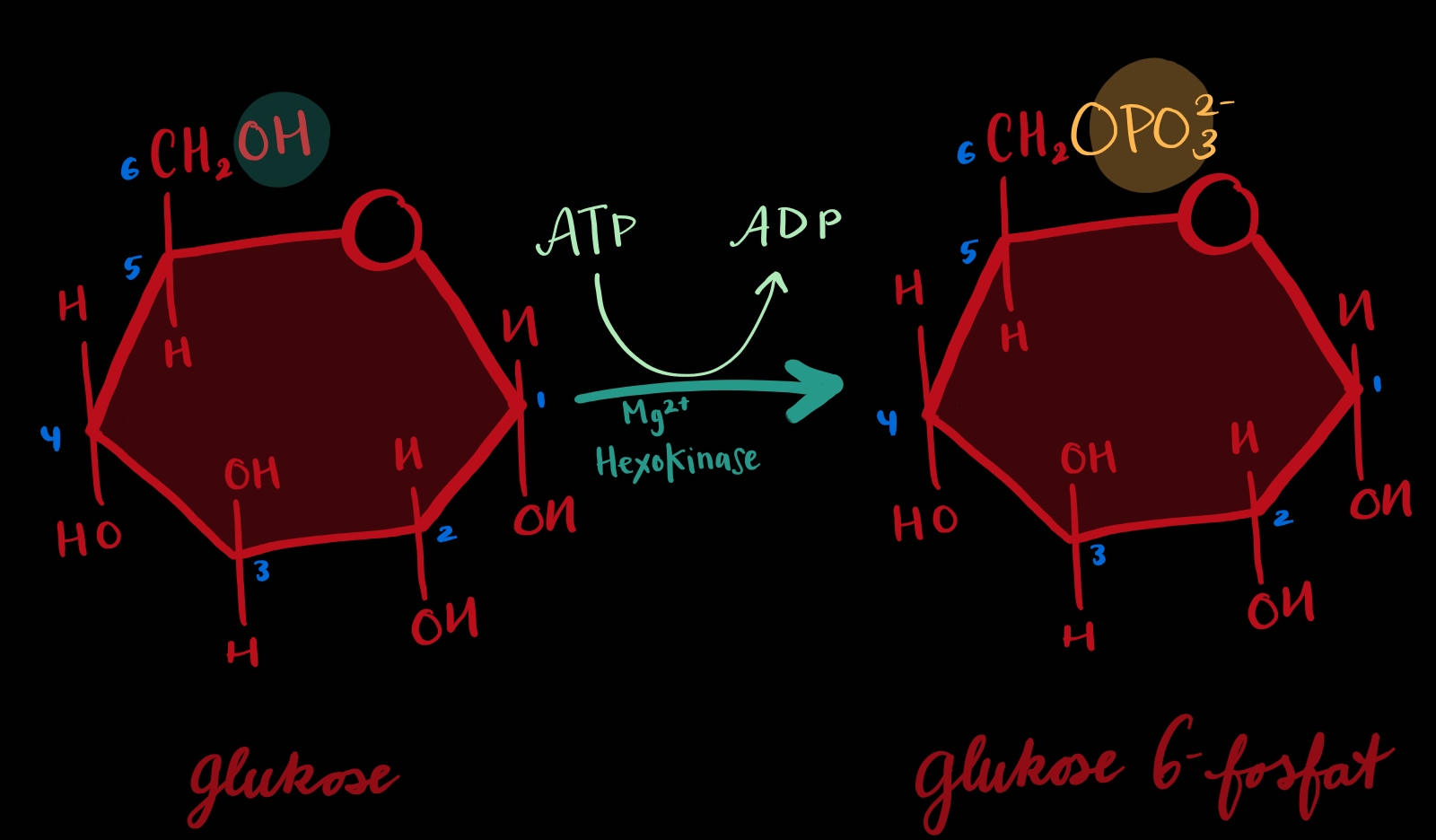
TRINN 1: Frå glukose til glukose 6-fosfat
Glukose blir fosforylert til glukose 6-fosfat.
Talet 6 betyr karbon nummer 6. Om ein ser molekylet teikna som ein ring begynner ein å telle karbon lengst oppe mot høgre og så med klokka (dette er min regel i alle fall 🧂). Karbon nummer 6 er altså oppe til venstre sida glukosen berre har 6 karbon. Og om det er eit karbon rett opp så er det det siste. Her betyr det at fosfatgruppa blir lagt til karbon nummer 6.
Å putte fosfat på molekylet er energikrevjande[1] og kan berre bli gjort saman med ein handling som gir frå seg engergi[2]. Om ein splittar den ytste fosfatgruppa frå eit ATP får ein frigitt mykje energi og det er det som blir gjort her. Naturlegvis er det det avsplitta fosfatet som blir tatt opp av molekylet.
Det er ei hydroksylgruppe[3] på molekylet som har ansvaret med å fange opp fosfatgruppa. Oksygenet vil binde seg til fosfatet og hydrogenet vil falle frå. Men fosfatet er svært negativt og det skremmer oksygenet vekk! 😱 Men det er her enzymet verkeleg kjem inn: enzymet har med seg mykje positive ion, Mg2+, som skjuler all den negative ladninga og viser berre fram fosfatets mest positive side til oksygenet, slik at oksygen og fosfat kan kople seg saman. No hjelper fosfatets negative ladning til å halde molekylet inni cella og utforbi alle organeller, plasmamembranen lar nemleg ikkje ladde molekyl få sleppe igjennom. Ja fordi glykolysen skjer i cytosol til forskjell frå resten av celleåndinga som skjer i mitokondriene.
TRINN 2: Frå glukose 6-fosfat til fruktose 6-fosfat
Molekylet blir gjort om frå glukose til fruktose. Dei har same formel, men ulik struktur. Formelen er C6H12O6. Om ein ser på strukturen, så ser ein at glukose har ein karbonyl[1] på fyrste karbonet mens fruktose har ein slik på andre karbonet. Denne forskjellen er òg grunnen til at glukose vert kalla aldose eller aldehyde, og fruktose vert kalla ketose eller ketone[2]. Denne dobbelbindinga er ikkje til stades når molekylet er i ring-form, men strukturen er framleis annleis sida ringen sluttar til eit anna karbon enn før, C5 slår seg i lag med C2 i staden for C1. Det betyr at glukosen lagar ein 6-kanta ring, mens fruktosen lagar ein 5-kanta ring (med ekstra vedheng).
Grunnen til at dette blir gjort er fordi ei karbonylgruppe ikkje kan fosoforylerast og me skal ende opp med ei fosfatgruppe i kvar ende. Molekylet skal jo delast på midten og bli to nokså like delar(og til slutt to heilt like delar). Eg må innrømme at eg ikkje veit kvifor, men det er det biokjemiboka mi seier.
Trinn 2 handlar altså om å flytte ein dobbelbinding ned to hakk (C1=O ➡️ C1=C2 ➡️ C2=O). Mellomtrinnet vert kalla ein enediol[3]. Prosessen er enkelt reversibel og ikkje særleg energikrevjande, men altså enzymkrevjande. Ein seier at molekylet blir isomerisert: at formelen er den same som før men strukturen har endra seg.
👩🏫
Det fyrste som ein må gjere er å opne opp ringen ✂️, så må elektrona manipulerast slik at bindingane flyttar på seg. Proton frå C2 blir plukka vekk og oksygenet på C1 blir freista av eit proton og gir opp si dobbelbinding med C1. C1 og C2 lagar dobbelbinding med kvarandre. No er molekylet midlertidig eit enediol.
Vidare blir det gjenværande protonet ved C2 plukka vekk og C2 dobbelbinder seg med oksygenet sitt i staden for C1 (ny karbonylgruppe og ketongruppe ved C2), og C1 bind seg til eit ekstra proton i staden. Og så lukkar ringen seg igjen og no har me fruktose 6-fosfat.
2: Eg forstår ikkje forskjell på aldose og alhyde, og ketose og ketone. Det føles som om orda blir brukt hipomhap 😕
3: det er ein dobbelbinding mellom to karbon der karbona har kvar si hydroksylgruppe (-OH)
TRINN 3: Frå fruktose 6-fosfat til fruktose 1,6-bisfosfat
Fruktosemolekylet får ei fosfatgruppe lagt til på fyrste karbonet slik at det har ei fosfatgruppe i kvar ende[1]. Dette er siste trinnet før molekylet blir splitta i to, og dette er det siste trinnet som bruker ATP i staden for å produsere ATP[2].
Om du lurer på kva som er forskjell på bisfosfat[3] og difosfat[4] så har bisfosfat fosfatgruppene festa på ulike stadar på molekylet, mens difosfat har fosfatgruppene festa etter kvarandre som ei lenke. For tre fosfatgrupper heiter det trisfosfat om gruppene er enkle og trifosfat om dei er festa i kvarandre.
Trinn 3 er eit skillepunkt for molekylet med tanke på framtida. Vanleg glukose, glukose 6-fosfat og fruktose 6-fosfat har alle saman andre moglegheiter i sine livsløp enn berre ein glykolyseprosess. Dei kan lyserast og dei kan ikkje. Men fruktose 1,6-bisfosfat blir berre brukt til glykolyse. Trinn 3 kan heller ikkje lett bli omgjort. Så et er altså eit skillepunkt for molekylet: utan glykolyse nå så blir det berre opphoping av fruktose 1,6-bisfosfat.
Akkurat som i trinn 1 så er det denne gong ein kinase som hjelper til her, men ikkje den same(phosphofructokinase). Men biokjemiboka legger til noko om enzymet og fosforyleringa som eg rett og slett ikkje forstår «Some bacteria and protists and perhaps all plants have a phosphofructokinase that uses pyrophosphate (PPi), not ATP, as the phosphoryl group donor in the synthesis of fructose 1,6-bisphosphate». Men eg finner ingenting om dette andre stadar? Noko som verker litt rart om det skal gjelde for alle planter?
Enzymet som blir brukt her i trinn 3 er svært påverkeleg. Biokjemiboka nemner fleire ting, men det som er mest forståeleg er at mykje ATP i cella slår AV enzymet og mykje ADP og AMP slår PÅ enzymet. Dette gir meining sida glykolysen produserer ATP, og om ein allereie har ein del er det vel ikkje behov for å arbeide ekstra.
Fotnoter:
1: Fostfatgruppe på C1 og fosfatgruppe på C6
2: glykolysen ender naturlegvis opp i pluss til slutt
3: fruktose 1,6 bisfosfat
4: adenosindifosfat (ADP)
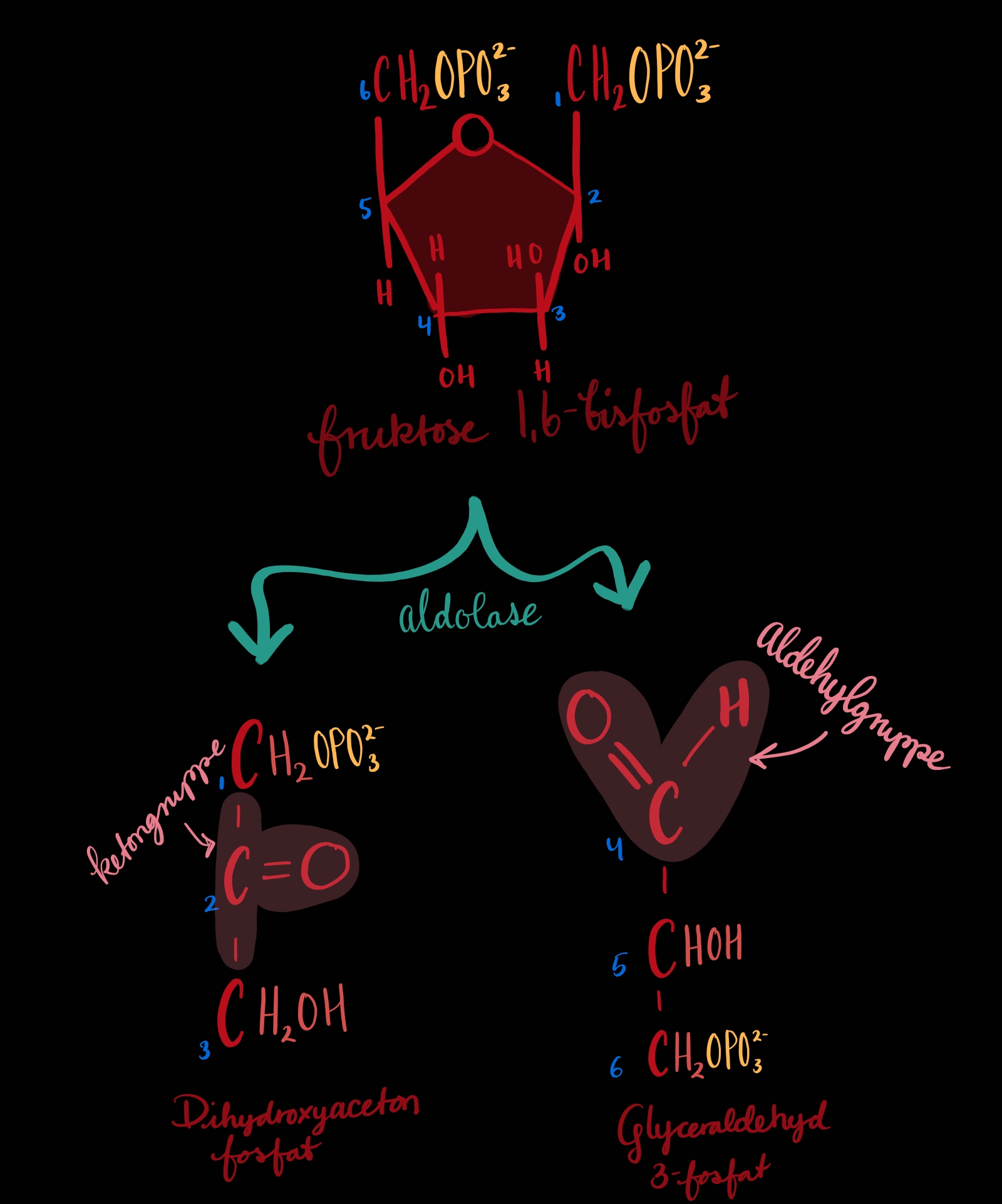
TRINN 4: Glykolysetrinnet - Sukkersplitting
Sukkeret, fruktose 1,6-bisfosfat, blir delt i to 💔. Begge delane har tre karbon kvar men dei er ikkje like i struktur. Glyceraldehyde 3-fosfat[1] er det den meir ideelle delen og kan gå rett til trinn 6. Dihydroksyaceton fosfat[2] har endå ikkje rett struktur og må gjennom trinn 5.
Biokjemiboka seier at glykolysen er avhengig av at karbonylet er på C2 (sjå trinn 2) for at splittinga skal skje på rett plass, mellom C3 og C4. Splittinga er ein aldolkondensasjon i revers. Eg er ikkje så god i dette så eg berre gjentar det boka seier. For min egen del så er det ein ganske detaljert illustrasjon på side 551 (mechanism figure 14-6).
Fotnoter:
1: eit aldehyd/aldose
2: eit ketone/ketose ... eg veit ikkje forskjell på desse fire orda (finst det fleire?) er det norsk/engelsk?, subjektiv/adjektiv? epler/appelsiner?
TRINN 5: Dihydroksyaceton fosfat blir til glyceraldehyd 3-fosfat
Dette er eit lite tilleggstrinn til trinn 4. Resultatet av trinn 4 var to ulike molekyl og me trenger to LIKE. Så ein må endre seg, og det er dihydroksyaceton fosfat. Dihydroksyaceton fosfat blir isomerisert - det var det som blei gjort trinn 2 òg[1], altså at molekylformelen består men strukturen skal endrast. Etter at dette er gjort er det ikkje mogleg å sjå forskjell på alle glyceraldehyd 3-fosfatane, om dei har ei fortid som dihydroksyaceton fosfat eller ikkje. Dei er alle prikkens like på alle måtar.
Dei siste trinna, trinn 6-10, blir teknisk sett gjort to gonger per glukosemolekyl som me starta med. Den opprinnelege glukosen er no delt i to ✌️, så det blir ein gjennomgong per halvdel. Som eg nevnte i introduksjonen er glukolysen betre illustrert som ein stemmegaffel enn ein vertikal strek.
Eg må innrømme eg veit ikkje nøyaktig korleis denne isomeriseringa foregår, men det er sikkert noko lignande som førre gong (trinn2) at proton blir flytta på og dobbelbindingen beveg på seg alt dit me vil ha han. Det blir på ein måte motsatt av trinn 2.
Fotnoter:
1: Då glukose blei til fruktose då gjekk me frå aldehyd til keton (frå karbonyl på enden til karbonyl på C2), denne gong er det motsett veg (karbonylet skal ende opp på enden)
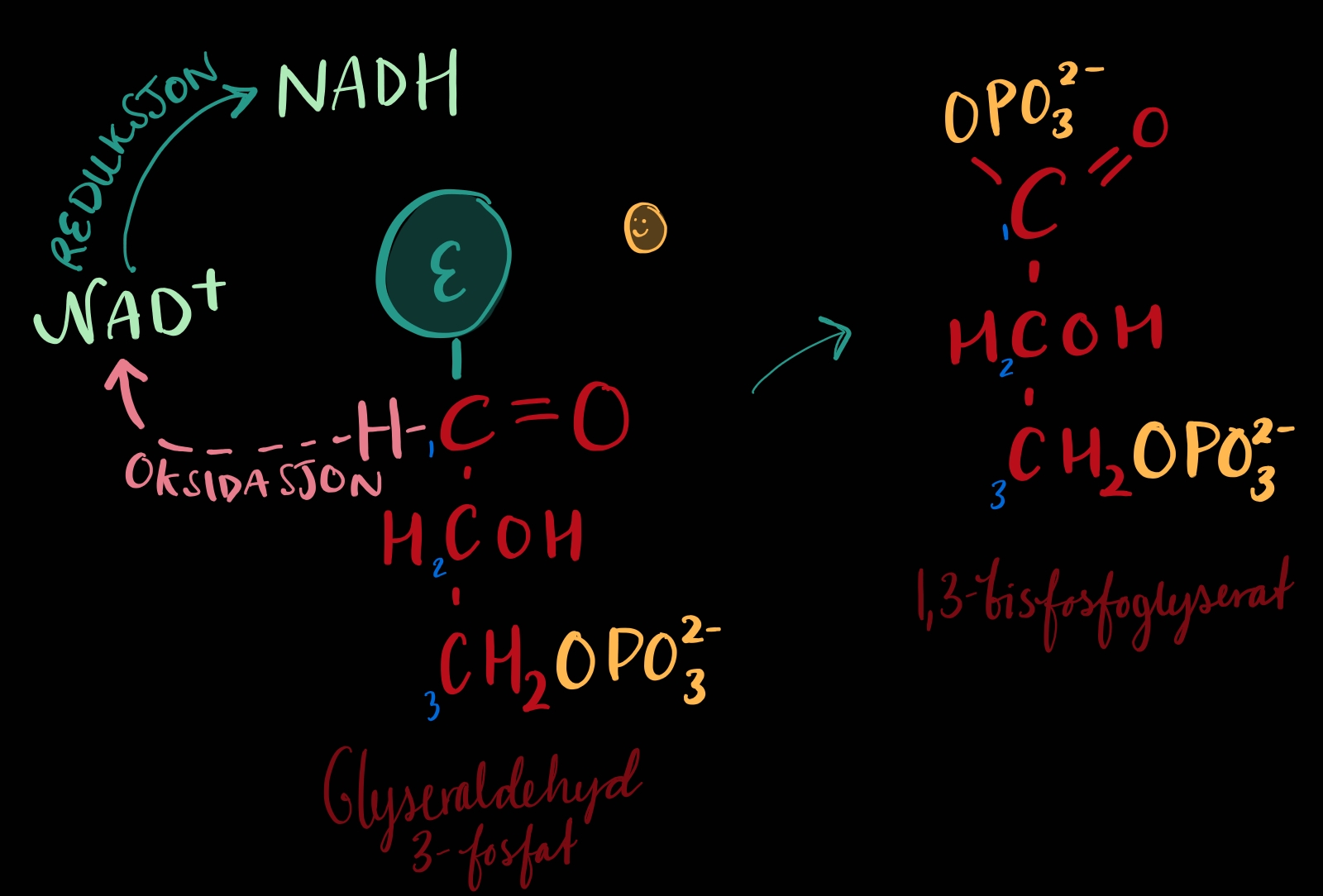
TRINN 6: Frå glyceralaldehyd 3-fosfat til 1,3-bisfosfoglycerat
Energiproduksjonen startar!
Trinn 6 kort oppsumert er at glyceraldehyd 3-fosfat blir oksidert[1] og så fosforylert. NAD+ blir redusert[2] til NADH. Oksidasjonen av molekylet, altså at hydrogenatomet drar frå molekylet, gir frå seg nok energi til å drive dei energikrevjande prosessane (1)formasjon av NADH og (2)formasjon av binding mellom molekylet og fosfatgruppa.
Me startar altså med glyceraldehyd 3-fosfatet, som har ei aldehydgruppe[3] i enden. Aldehydgruppa bind seg til enzymet og til eit NAD+ som finst der. Glyceraldehyd 3-fosfat mister eit hydrogenatom og eit elektron(oksidasjon) til NAD+ som reduserast til NADH. Oksidasjonen gir frå seg ein del energi, noko blir lagra i NADH-et og noko blir lagra i ein høgenergibinding mellom vårt molekyl og enzymet. No kjem eit uorganisk fosfat Pi (Dette er ikkje ei fosfatgruppe frå noko ATP) og koplar seg til molekylet, eller fosforylerer molekylet, der enzymet til no har vore bunden. Framleis har me ei høgengergibinding mellom fosfatet og oksygenet som koplar fosfatgruppa til molekylet - om eg har forstått det riktig. I neste trinn vil i alle fall hydrolysen/oppløysinga av dette høgenergibandet vere viktig (kor enn det er).
No heiter vårt nye molekyl 1,3-bisfosfoglycerat. Me hadde fosfat på C3 me oss frå før og fekk no eit nytt fosfat på C1.
PS: Teikningane mine viser for så vidt det same men på litt ulike måtar.
Fotnoter:
1: Oksidasjon er tap av elektron
2: Reduksjon er at ein får elektron
3: karbonet på enden med dobbelbinding til oksygenet og enkeltbinding til hydrogenet er ei aldehydgruppe
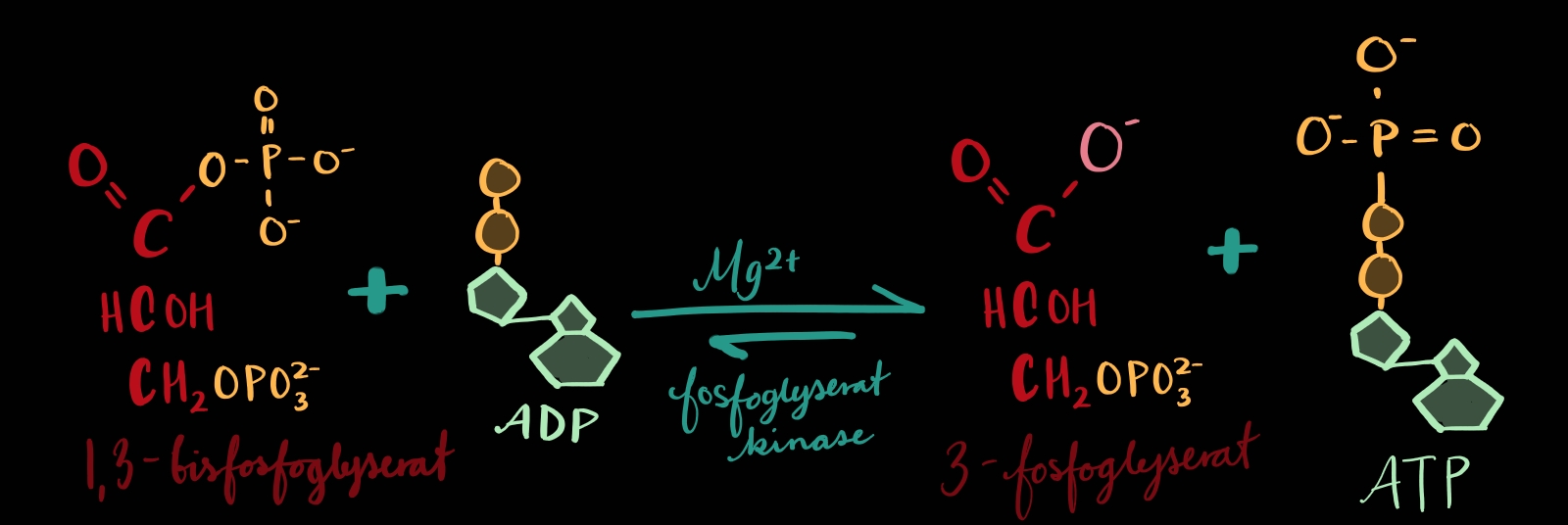
TRINN 7: Frå 1,3-bisfosfoglyserat til 3-fosfoglyserat
Energien som blei lagra på molekylet i førre trinn blir nå gjort til nytte: I trinn 6 blei det lagt til ei fosfatgruppe på C1 og ho* blir no overført til eit ADP(som blir til eit ATP).
*Om du ser på teikningen min så er det faktisk ikkje heile fosfatgruppa som blir overført til ADP-et, eit oksygen blir igjen. Derfor er det ikkje eit fosfat det er snakk om i trinn 7, men ei fosforyl-gruppe. Så vidt eg forstår er det akkurat det same, bortsett frå eit oksygen. Det gir meining nok sida det er ikkje plass til fire oksygen om gruppa skal kople seg på eit fullt oksygen ytterst på ADP.
I førre trinn nemnte eg at me hadde med oss ei høgenergibinding[1] som ville vere viktig her i trinn 7. Om bindinga blir hydrolysert[2] så vil nemleg nok energi bli frigjort til å putte fosfatet frå trinn6 på eit ADP(som gjer oss ATP). Bindinga er om eg har forstått riktig: mellom fosforet(P) og oksygenet som bind fosfatgruppa til vårt molekyl. Og som sagt så gjort, bindinga blir hydrolysert og eit ADP får ei fosforylgruppe og blir eit ATP.
Trinn 6 og 7 heng altså nøye saman: fyrst blir det lagt til eit ikkje-ATP-fosfat og så blir fosfatet hekta på eit ADP som blir til eit ATP i trinn 7. Og i dette ATP-et er det lagra energi i bindingane som kan brukast til andre ting.
Fotnoter:
1: I trinn 6 blei det laga ei høgenergibinding mellom molekylet og enzymet (tioesterband) og dette bei overført til fosfatgruppa då ho blanda seg inn(fosfatband)
2: Splitting av eit kovalentband(når atoma deler elektron t.d. H:H) vha vatn. Altså eine produktet får -H og hin får -OH
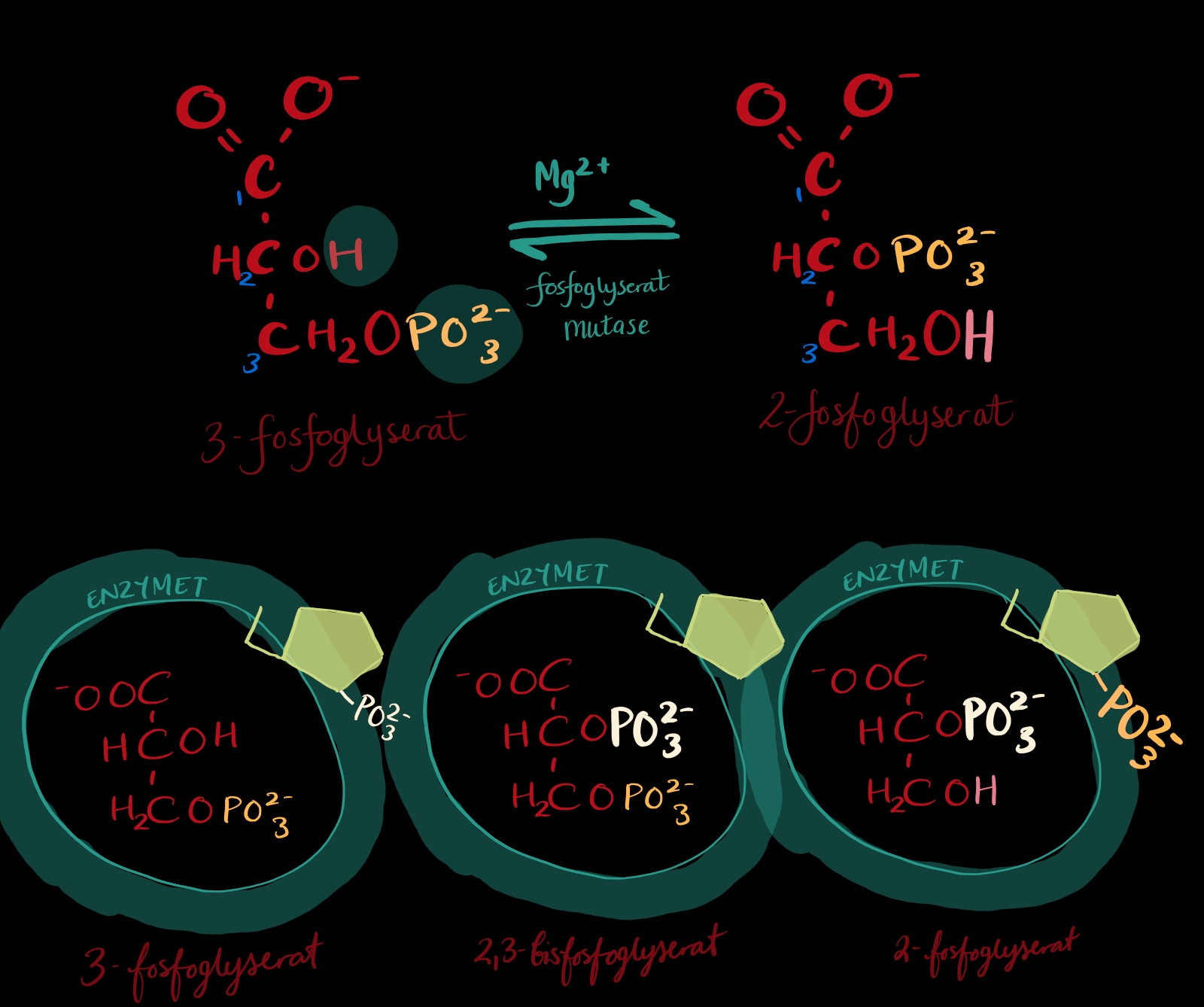
TRINN 8: Frå 3-fosfoglyserat til 2-fosfoglyserat
Den gjenverande fosfatgruppa frigjer relativt lite energi på C3 og er betre tent med å vere på C2. Nå trur du kanskje at ho berre flyttar seg opp eitt hakk? Det ser nemleg slik ut på teikningar, men det er litt misvisande.
Fyrst, så er det snakk om fosforylgrupper akkurat som i trinn 7, dvs. eit fosfor med 3 oksygen i staden for 4, ingen stor deal. For det andre så er det snakk om to fosforylgrupper: det bli lagt til ei fosforylgruppe på C2 før fosforylgruppa på C3 drar, altså så har molekylet ei bittelita stund på trinn 8 to fosfatgrupper på seg. Men så forsvinn fosforylet på C3 og me står igjen med 2-fosfoglyserat.
Når ein ser på teikningane og leser ei skildring av kva som skjer på trinn8 så minner det litt om trinn2 og 5, desse trinna var isomeriseringar og brukte enzym som slutta på isomerase. I trinn 8 heiter enzymet mutase og det er IKKJE skildra som ein isomerisering. Eg lurar på om det er fordi molekylet denne gong ikkje stokkar om på det det allereie har, men puttar på ei heilt ny gruppe som kjem utanfrå. Elles ser eg ikkje store forskjellen.
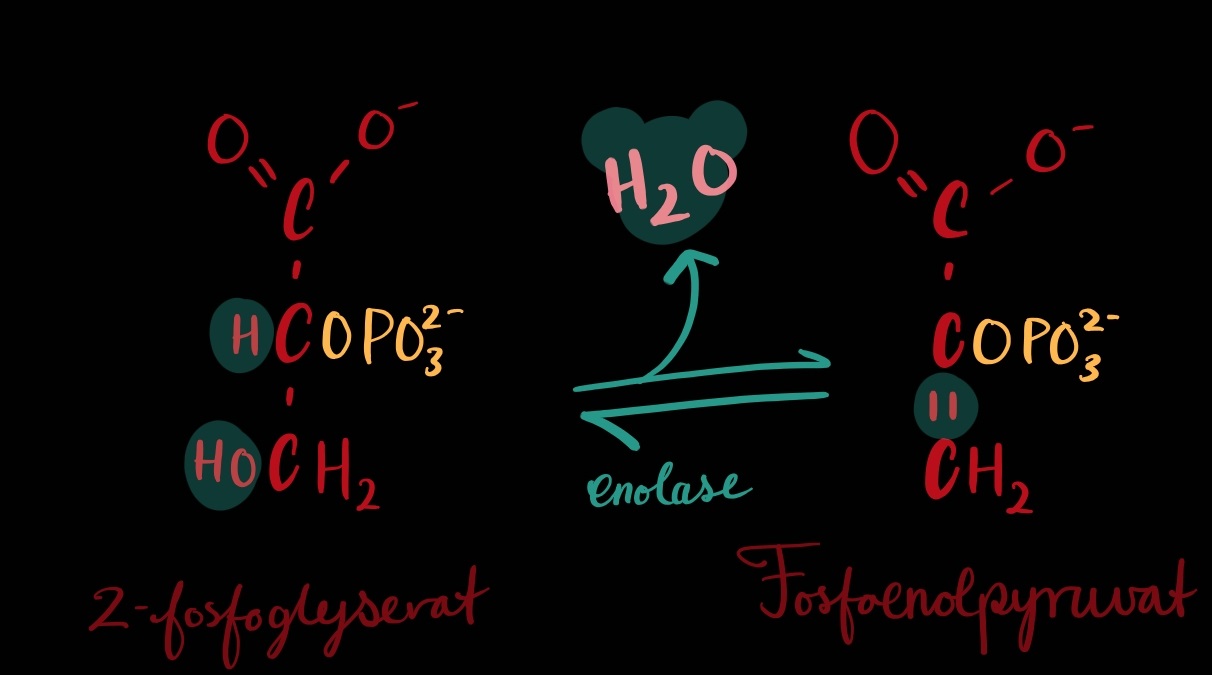
TRINN 9: Frå 2-fosfoglyserat til fosfoenolpyruvat
Detta trinnet likner på ein måte på trinn 6. Det handlar om å lage ein høgenergiforbinding slik at ein kan hausta energien i neste trinn.
Vatn blir fjerna[1] og eig høgenergi-enol-fosfat-band blir forma, seier cellebiologiboka – eg forstår ikkje kva det heilt betyr og slit med å forstå kor enolet er på vårt molekyl?[2]. Men eg får berre godta at slik er det.
Fotnoter:
1: Dehydrering
2: Eit enol er så vidt eg forstår to karbon med dobbelbinding og eine karbonet har eit hydroksyl (-OH)
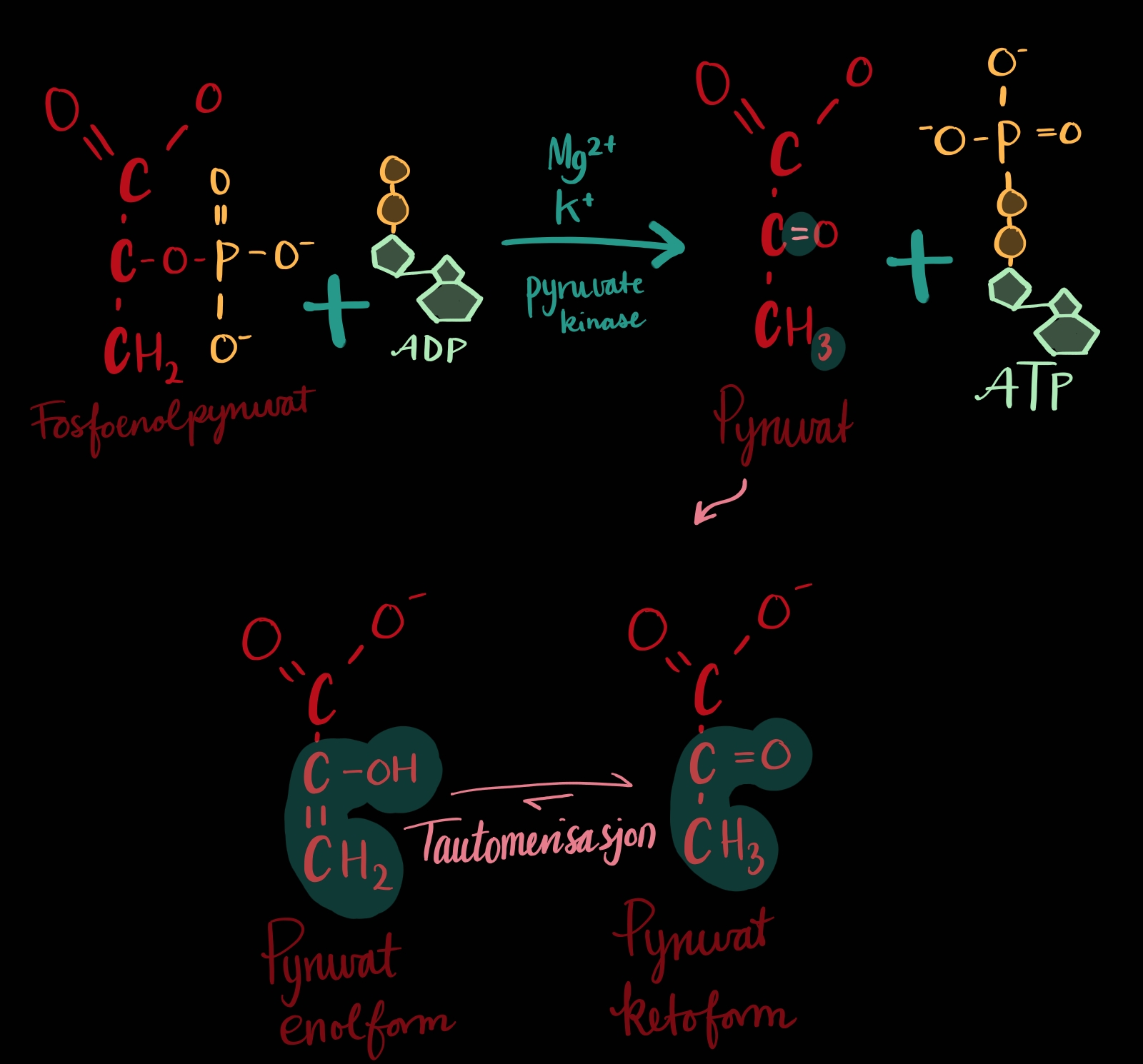
TRINN 10: Frå fosfoenolpyruvat til pyruvat
No, som i trinn 7, så blir det overført ei fosforylgruppe frå vårt molekyl til eit ADP slik at me får eit ATP, sprekkferdig med energi og eit pyruvat. Ja altså, to pyruvat for kvart glukose som me starta med - trinn 6,7,8,9 og 10 blir jo gjort to gonger per glukose fordi molekylet blei delt i to på trinn 4.
Rett etter at fosforylgruppa har dratt så vil pyruvatet tautomerisere[1] seg, frå enol-form til keto-form[2]. Dette trengs ikkje enzym for, han gjer det sjølv.
Fotnoter:
1: Molekylet kan endre raskt sin egen struktur alt etter som kva dei vil vere, eg trur det er avhengig av pH?
2: Eg trur dette berre er at proton kjem og går slik at dobbelbindingen flytter seg alt etter som kor det er plass
Kjelder
Alberts, B., Hopkin, K., Johnson, A. D., Morgan, D., Raff, M., Roberts, K. & Walter, P. (2014) Essential Cell Biology (4. utg.) Taylor & Francis Group
Evert, R. F & Eichhorn, S. E., (2013) Raven Biology of Plants (8. utg.) W. H. Freeman and Company
Nelson, David L. & Cox, Michael M. (2013) Lehninger Principles of Biochemistry (6. utg.) W. H. Freeman and Company
Widmaier, E. P., Raff, H., Strang & Strang, K. T.,(2011) Vander's Human Physiology (12. utg.) McGraw-Hill